 Line drawing of male plant with flowers by M. Szent Ivany, J. Adelaide Bot. Gards 4 (1981) 299, fig. 139. |
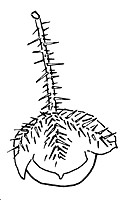 Sketch of fruit by D.E.Symon, J. Adelaide Bot. Gards 4 (1981) 341, fig. 158F |
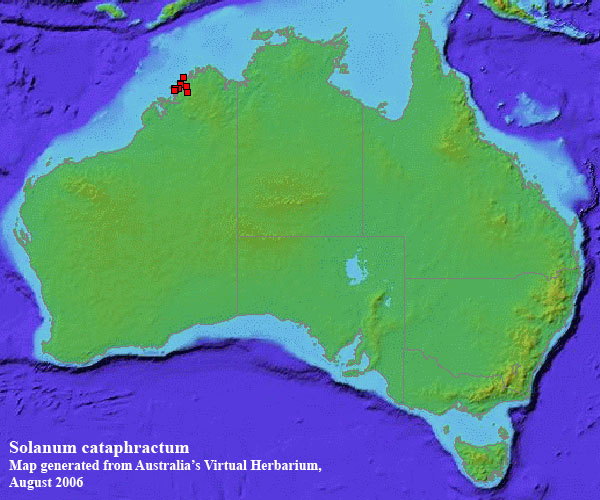 Distribution map generated from Australia's Virtual Herbarium. |
Synonymy
Solanum cataphractum Cunn. ex Benth., Fl. Austral. 4: 459 (1868)
T: (Prince) Regent River, W.A., A. Cunningham 132; lecto: BM; iso: K p.p., MEL; Bat Island, W.A., A. Cunningham 192; syn: BM p.p. K, p.p., fide D. E. Symon, J. Adelaide Bot. Gard. 4: 298-300 (1981).
Images of the lectotype sheet and another syntype can be seen on the Solanaceae Source site.
Description
Erect or sprawling shrub, green, male or female; younger parts sparsely pubescent with minute stellate hairs, the stems and leaves soon glabrous; prickles to 7 mm long, abundant on all parts.
Leaves lanceolate; lamina 3-6 cm long, concolorous, deeply lobed; lobes narrowly linear; petiole c. 1 cm long.
Male plant: inflorescence 6-20-flowered; rachis to 8 cm long; pedicels c. 5 mm long; calyx 5-7 mm long, the lobes lanceolate, 3-4 mm long; corolla rotate, 20 mm diam., purple; anthers 3 mm long. Female plant: flowers solitary, pedicels c. 8 mm long, calyx 7-9 mm long, enlarged in fruit, the lobes linear, 4-5 mm long; anthers 2-5 mm long. Berry c. 20 mm diam.
Distribution and ecology
A poorly known species, restricted to the coast and adjacent offshore islands of the Kimberley, W.A.
Relationships
All of the dioecious members of the Dioicum group of subgen. Leptostemonum exhibit functional dioecy i.e. male flowers are borne on separate plants from what appear to be plants with bisexual or hermaphrodite flowers; while these bisexual flowers do produce pollen, it is inaperturate and does not germinate, making the flowers effectively female (Knapp et al.,1998).
Initial molecular studies by Martine et al. (2006) suggested that the two functionally dioecious species from Kakadu, S. asymmetriphyllum and the newly described S. sejunctum, formed a separate clade and had arisen separately from the other dioecious members of the Dioicum group, S. cunninghamii, S. cataphractum, S. dioicum, S. petraeum, S. carduiforme, S. tudununggae, S. vansittartense and S. leopoldens. T his initial DNA analysis involved sequencing of the ITS region of nuclear rDNA.
However further molecular analysis involving the trnK-matK gene region indicated that all of the functionally dioecious species form a single clade arising from the andromononoecious species of the group (Martine et al., 2009).
Many of these dioecious species remain undercollected and poorly understood and it is likely that some of these names will be changed with further work. S. cataphractum was not part of the second molecular study and only a single specimen was included in the first study (Martine et al., 2006).
References: S.Knapp, V.Persson & S.Blackmore (1998). Pollen morphology and functional dioecy in Solanum (Solanaceae). Pl. Syst. Evol. 210:113-139.; Martine, C.T., D. Vanderpool, G.J. Anderson, and D.H. Les (2006). Phylogenetic relationships of andromonoecious and dioecious Australian species of Solanum subgenus Leptostemonum section Melongena: Inferences from ITS sequence data. Systematic Botany 31: 410-420; Martine, C.T., G.J. Anderson & D.H. Les (2006). Gender-bending aubergines; molecular phylogenetics of cryptically dioecious Solanum in Australia. Australian Systematic Botany 22: 107-120.
Selected specimens
W.A.: Bigge Is., N.G. Marchant 72/42 (PERTH); Sandstone country where G. Grey had his main base camp in 1837, 1522’04” 12445’19”, A.A.Mitchell 3469 (AD, BROOME, PERTH).
Plant status, if any
Originally classified as Risk Code 2K (Briggs & Leigh, Rare or Threatened Australian Plants, 1995) but this species is no longer considered to be at risk - see FloraBase
From the web
Further information for this species in WA can be found on the FloraBase site.