 Illustration by G. Dashorst to accompany protologue. LHS plant is bisexual or fruiting plant; RHS is male plant. |
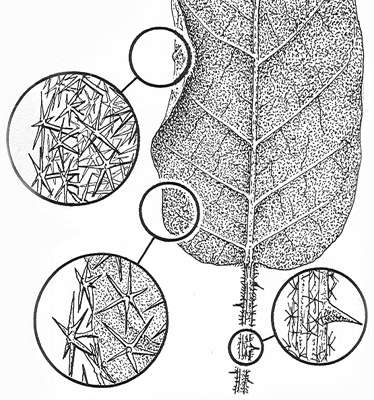 Detail of leaf hairs from upper and lower leaf surface and petiole. From illustration by G.Dashorst accompanying protologue. |
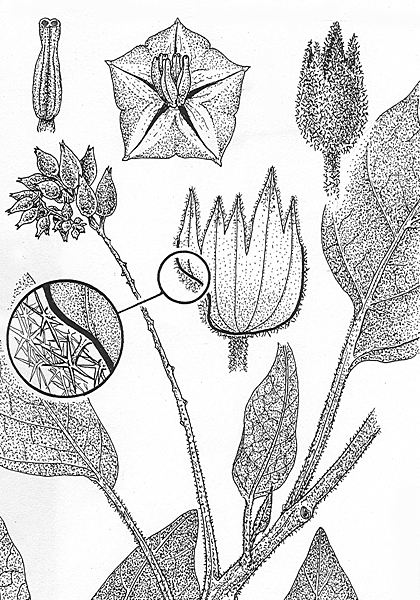 Detail of male inflorescence and flowers. From illustration by G.Dashorst accompanying protologue. |
 Detail of fruit within enclosing prickly calyx and seed. From illustration by G.Dashorst accompanying protologue. |
Synonymy
Solanum sejunctum K.Brennan, C.Martine & Symon, Beagle 22: 1-7.
T: NT, N facing wall in central part of Mt Brockman, 23 Feb 1973, L.A.Craven 2371; holo: DNA; iso: CANB.
Solanum A55445 "Mt Brockman" and Solanum sp. Mt Brockman (L.A.Craven 2371): Checklists of NT Vascular Plants (October 2005 and prior).
Description
Erect, clonal shrub to 1 m, male or female, grey-green, pubescent with stellate hairs; prickles to 3 mm long, common on calyx of bisexual flower, sparse to absent elsewhere.
Leaves elliptic to ovate-elliptic; lamina 5-11 cm long, 2.5-5.5 cm wide, concolorous by pubescence on both surfaces, entire; petiole 1-2.5 cm long.
Male plant: inflorescence elongate, up to 40-flowered; peduncle to 11 cm long; pedicels 5-8 mm long; calyx c. 6 mm long, the lobes shortly acuminate; corolla broadly stellate or rotate-pentagonal, 20-30 mm diam., purple; anthers c. 5 mm long. Fruiting plant: flowers solitary, larger than male flowers; pedicel length unknown, but c. 7 mm long in fruit; calyx enlarged in fruit, the lobes narrowly triangular with linear apices, 5-10 mm long; corolla rotate, ?30-40 mm diam., purple-blue, lobes with acumens; anther length unknown.
Fruiting calyx c. 3 cm diam. Berry globular, 2-3 cm diam., ?green when ripe, drying black. Seeds c. 2.5 mm long.
The bisexual flower was known only from a photograph when this species was described. Consequently there is missing information in this description, compiled from an early draft of the manuscript [rmb].
Distribution and ecology
Primarily known from the Arnhem Land escarpment in Kakadu National Park, particularly the Mt Brockman and Nourlangie areas.
Occurs in sandstone in Allosyncarpia forest.
Relationships
All of the dioecious members of the Dioicum group of subgen. Leptostemonum exhibit functional dioecy i.e. male flowers are borne on separate plants from what appear to be plants with bisexual or hermaphrodite flowers; while these bisexual flowers do produce pollen, it is inaperturate and does not germinate, making the flowers effectively female (Knapp et al.,1998).
Initial molecular studies by Martine et al. (2006) suggested that the two functionally dioecious species, S. asymmetriphyllum and the newly described S. sejunctum, both from Kakadu, formed a separate clade and had arisen separately from the other members of the Dioicum group.
Three collections of S. asymmetriphyllum and three of S. sejunctum were included in this initial DNA analysis involving sequencing of the ITS region of nuclear rDNA. However further molecular analysis involving the trnK-matK gene region indicated that these dioecious species should all form a single dioecious clade arising from the andromononoecious species of the group (Martine et al., 2009).
References: S.Knapp, V.Persson & S.Blackmore (1998). Pollen morphology and functional dioecy in Solanum (Solanaceae). Pl. Syst. Evol. 210:113-139.; Martine, C.T., D. Vanderpool, G.J. Anderson, and D.H. Les (2006). Phylogenetic relationships of andromonoecious and dioecious Australian species of Solanum subgenus Leptostemonum section Melongena: Inferences from ITS sequence data. Systematic Botany 31: 410-420; Martine, C.T., G.J. Anderson & D.H. Les (2006). Gender-bending aubergines; molecular phylogenetics of cryptically dioecious Solanum in Australia. Australian Systematic Botany 22: 107-120.
Notes
S. sejunctum can be distinguished from S. asymmetriphyllum by its more or less concolorous leaves; leaves of S. sejunctum are pubescent on both surfaces whereas they tend to be glabrous on the upper surface and densely pubescent on the lower surface in S. asymmetriphyllum and consequently the leaves are usually somewhat discolorous.
Prickles on the calyces of S. asymmetriphyllum are up to 10 mm long whereas they are only ever up to 2 mm long in S. sejunctum.
S. asymmetriphyllum has been recorded with shortly lobed leaves, whereas they have only ever been recorded as entire in S. sejunctum to this time.
Plant status, if any
Conservation status as a Data Deficient plant (as Solanum sp. Mt Brockman) in the
From the web